Cèl.lules mare: Un nou concepte en medicina regenerativa
M.Eugenia Mato Matute 1,2
1 Fundació Sardà Farriol, 2 Departament de Biologia Cel.lular i Anatomia Patològica, Facultat de Medicina. Universitat de Barcelona,
Durant la vida d'un organisme, les cèl•lules que formen part dels teixits, siguin de manera natural o bé per una malaltia (aguda o crónica), pateixen un desgast que condueix a la seva degeneració. Els avenços en la medicina basada en tècniques substitutories del teixit danyat ha suposat una revolució no exempta de problemes, entre els que cal destacar tant la limitació en el nombre de donació d'òrgans, com de les complicacions immunològiques (rebuig de l'empelt), complicacions aquestes últimes pal•liades en part de manera farmacològica.
De manera natural, els teixits que formen part del nostre organisme tenen la capacitat intrínseca d'autorenovar-se, fet que es produeix gràcies a cèl•lules que anomanem mare o troncals amb la seva capacitat de diferenciació. Aquest fenomen ha obert una nova era en l´anomenada medicina regenerativa, que no és més que aprofitar els mecanismes naturals de renovació cel•lular, per poder reparar els teixits danyats. Sens dubte que aquest nou concepte en la medicina no només ha obert noves possibles vies terapèutiques d'estudi, teràpies cel•lulars, sino ha obert la caixa de Pandora per qüestions no exclusivament científiques que exigeisen un debat ètic, per part de la societat.
Definició i tipus de cèl•lula mare: Es defineix com una cèl•lula totipotent/pluripotent o multipotent capaç de generar un o més tipus de cèl•lules diferenciades i que a més posseeix la capacitat de autorenavació ,és a dir de produir més cèl•lules mare.
En els animals superiors, les cèl•lules mare u troncals es classifiquen en els següents grups:
1.- Cèl•lules mare embriòniques :
1.1.- Cèl.lules mare embriòniques o (ES)
1.2.-Cèl•lules mare germinals (EG)
1.3.-Cèl•lules mare dels teratomes i teratocarcinomes1.1.- Cèl.lules mare embriòniques o (ES). Deriven de la massa cel•lular interna de l'embrió en l'estadi de blastòcits (7-14 dies), consideradas com a totipotents /pluritopents. A partir d'elles, i després de moltes divisions cel•lulars, esdevindran cèl.lules diferenciades, que formaran part del teixit especialitzat. Malgrat tot, i que encara les cèl•lules de la massa cel•lular interna del blàstocit sòn pluripotents, no són per si mateixes cèl•lules mare dins de l'embrió, perquè aquestes no es mantenen indefinidament com a tals en condicions in vivo, sinó que es diferencien successivament en els diversos tipus cel•lulars durant la fase intrauterina. El que succeeix és que quan s'extreuen de l'embrió i es mantenen sota certes condicions in vitro, aquestes es converteixen en cèl•lules “immortals” dotades de dues propietats: autorenovació i pluripotència. Característiques importants per a poder ser utilitzades en teràpia cel•lular.
1.2.-Cèl•lules mare germinals (EG): Es localitzen en la cresta germinal dels fetus, lloc on es produeix la diferenciació de la línia germinal.
1.3.-Cèl•lules mare dels teratomes i teratocarcinomes: Es localitzen en les gònades en forma de tumoració. Les cèl•lules diferenciades del tumor es formen a partir de cèl•lules mare pluripotents de carcinoma embrionari que deriven a la seva vegada de cèl•lules primordials germinals de l'embrió (post-implantació). Són tumors que contenen una gran varietat de tipus cel•lulars que inclouen des de cèl•lules musculars, cartílag, os, epiteli, neuroextoderm primitiu, estructures ganglionars i epiteli glandular, és a dir estirp cel.lular derivades de les tres capes embrionàries que té un embrió (endoderm, mesoderm ectoderm ). Aquestes cèl•lules diferenciades del tumor mantenen la capacitat pluripotencial quan son mantingudes en condicions in vitro. Un dels experiments més espectaculars va ser que després del tractament d'una línia cel•lular d'un tumor testicular amb àcid retinoic, aquestes cèl-lules es diferenciesin a cèl•lules nervioses. (1-6).
(Fig.1)
2.- Cèl.lules mares o òrgan-específiques (adult)
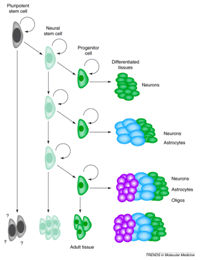
Derivades de les embriòniques, mantenen al llarg de la vida del teixit la capacitat multipotencial, és a dir són capaces d'originar cèl•lules especialitzades d'un òrgan concret no només en l'embrió, sinó també en l'adult. L'exemple més clar d'aquest tipus cel•lular és el de les cèl•lules de la medul•la òssia, que són capaces de generar tots els tipus cel•lulars de la sang i del sistema immune (7). Encara que es conèixen des de fa temps l' existència d'aquestes cèl•lules en els diferents teixits, en els últims anys diferents autors han identificat cèl.lules mare que provenen de la medul•la òssia i que, com a la sang o la epidermis presenten una gran taxa de proliferació. Presentan algunes d'elles presenten la suficient flexibilitat o plasticitat com per a generar cèl•lules especialitzades d'altres llinatges no hematològic (fenomen de trandiferenciació) (8-9). Això ha suposat una sorpresa encoratjadora, ja que augmenta la perspectiva d'obtenir a llarg i mig termini teràpies cel•lulars, sense els problemes ètics que comporta la destrucció d'embrions congelats. La pregunta clau és com, quan i el per què una cèl•lules mare en un teixit adult comencen a diferenciar-se. Existeixen múltiples mecanismes dins del “nínxol “ o hàbitat cel•lular, que produeixen senyals internes i externes. Aquest conjunts de senyals, permeten que es produeixen divisions simètriques i asimètriques, que donaran lloc al manteniment de les pròpies cèl•lules mare, com l'inici de la diferenciació cel•lular. Això es produeix, a partir de factors de transcripció que actuen en gens específics. (Fig.2)
Investigacións bàsicas en Drosophila melanogaster ( mosca del vinagre) i en ratolí han demostrat l'existència d'una sèrie de controls interns que a semblança de rellotges interns, marquen a les cèl•lules el nombre de divisions simètriques abans de començar el procés de diferenciació que donarà lloc a un llinatge cel•lular concreta. A pesar dels pocs coneixements que es tenen d'aquests processos, els estudis bàsics indiquen d'una banda l'existència de proteïnes específiques (activadores/inhibidores) implicades en el cicle cel•lular, així com el paper important que juguen la longitud del telòmer.
Definim el telòmer (del grec telos: final i mers: part) com els extrems dels cromosomes format per regions repetides d’ADN no codificants, amb funcions molt concretes en l'estabilitat molecular. El nombre de repeticions que es troben en el telòmer és variable, fins i tot en diferents cèl•lules d'un mateix individu, encara que el terme mitjà tendeix a ser constant per a cada espècie. D'altra banda, l’ enzim telomerasa (formada per un complex proteïna-àcid) és una polimerasa produïda per les cèl•lules germinals o embriòniques que permet l'elongació dels telòmers. És conegut que la telomerasa és reprimida en les cèl•lules somátiques madures, el que comporta un escurçament del telòmer al final de cada divisió cel•lular. Quan la longitud del telòmer arriba a cert límit, s'interrompen les mitosis quedant les cèl•lules en estadi Go del seu cicle (10). També es pot parlar de controls externs: un conjunt de senyals paracrines i autocrines entre les cèl•lules mare/filles i les cèl•lules veïnes, entre elles cal destacar:
a) Factors secretats per les cèl.lules: els millor coneguts són els factors de la medul•la òssia entre els quals s'inclouen nombroses citoquines ,
b) Interacció cèl•lula-cèl•lula a través de proteïnes de membrana,
c) Integracions de les cèl•lules amb la matriu extracel.lular per mitjà de receptors especìfics de membrana cel.lular: integrinas.
En molts d'aquests casos, el senyal extern interacciona amb el receptor de membrana i això, es tradueix cap a l'interior mitjançant una cascada de senyalitzacions, que no és més que un conjunt de reaccions bioquímiques: fosforilacions i desfosforilacions de proteïnes, per a finalment acabar en l'activació i desactivació d' un grup de gens. Això implica que la cèl•lula pot canviar el seu patró d'expressió, el que en determinats casos significa un pas més en la seva ruta de diferenciació, la seva proliferació, o fins i tot pot significar la mort cel•lular programada o apoptosi. Existint un delicat equilibri entre aquests processos cel•lulars.
PLASTICITAT DE LES CÈL•LULES MARE: CONCEPTE DE TRANSDIFERENCIACIÓ CEL•LULAR
Investigacions actuals donen una major evidència que les cèl•lules mare adultes tenen la capacitat pluripotencial en resposta a un estrès tisular. Existeixen nombrosos treballs científics que demostren la colonització per part de cèl•lules mare circulants a la sang perifèrica d'òrgans sotmesos dany tisular, tenen la capacitat de regenerar/reparar el teixit danyat. No obstant això; els resultats han de ser acuradament analitzats a pesar que existixen claras evidències d'aquests processos (11-13). En el procés de transdiferenciació es requereix que una cèl•lula posi en marcha mecanismes de desdiferenciació i re-diferenciació que donaran com a punt final altre tipus cel•lular.
En els últims anys, un nou concepte ha sorgit i és el de la fusió cel•lular. Aquest fenomen ha estat recentment demostrat en experiments portats a terme amb co-conreu de cèl•lules procedents de la medul•la òssia amb cèl•lules del cor, del fetge o cèl•lules neuronals. La fusió entre aquestes cèl•lules provoca un aportació de material genètic nou que podria permetre la substitució de les cèl•lules que s'estan degenerant per cèl•lules noves. No obstant això, aquestes desdiferenciacions aparents van resultar amb el producte de cèl•lules híbrides (tetraploids) (14). Aquestes aberracions cromosòmiques impedeixen la seva utilització en algun tipus de teràpia cel•lular. D'altra banda, existeixen treballs que demostren que els conreus de cèl•lules mare podrien produir tant canvis en la cinètica del cicle cel•lular, com a fenòmens d’adhesió cel•lular. (15-16)
EL SISTEMA IMMUNE ENFRONT DEL TRASPLANTAMENT DE CÈL•LULES MARE
Un dels problemes que planteja les teràpies del transplantament són els immunològics que acaben amb el rebuig de l'empelt. No obstant això, la teràpia alternativa d'obtenció, expansió i diferenciació de les cèl•lules mare requereix en ùltima instancia el transplantament de les mateixes, amb el risc evident d'un rebuig d'aquest empelt cel•lular. Malgrat tenir abundant experiència clínica, no es poden extrapolar els resultats de les dues tècniques, ja que per exemple en el cas de les cèl•lules no posseeixen vascularització.
El pròces de rebuig s’inicia amb un reconeixement del empelt per part dels limfòcits T-citotòxics a molècules del complex d’histocompatibilitat major MHC de classe I i classe II, expressades en la superfície cel•lular. Recentment s'ha demostrat nivells baixos d'expressió d'aquestes molècules en cèl•lules mare embrionàries humanes (17,18), i encara que augmenten al diferenciar-les in vitro no arriben mai als nivells dels observats en òrgans sencers. Degut a la poca o nul•la experiència in vivo, poc es coneix del poder antigènic d'aquestes cèl•lules, si bé hi ha estudis on descriuen nivells baixos i inducció de resposta immunològica. Sense oblidar-nos de les cèl•lules “natural killer” que intervenen de forma activa en la cascada de reaccións que condueixen al rebuig. No s'ha observat l'expressió del MHC de classe II en les cèl•lules mare (18).
Per aquest motiu, existeixen projectes de creació de bancs de cèl.lules mare isotipades a semblança dels bancs d´òrgans on es puguin escollir l´isotip més adient al pacient.
DIABETES MELLITUS I CÈL•LULES MARES
La Diabetis Mellitus pot subdividir-se en dos grans malalties: la Diabetis Mellitus tipus I (insulino-dependent) o DM1 caracteriztada per un procés autoimmune de destrucció de les cèl.lules productores d’insulina, el que provoca la falta d’aquesta hormona, i la Diabetis Mellitus tipus II (no insulino-dependent) que representa un 90% dels casos diagnosticats , la seva aparició es deu a la combinació d’una resistència a l’acció de la insulina per part dels teixits perifèrics i d’una alteració de la funció de la cèl.lula ß pancreàtica. Aquesta disfunció sembla ser el resultat de la incapacitat de la cèl.lula ß per produir i secretar insulina quan augmenta la demanda de la mateixa. La Diabetis afecta a un 4-5 % de la població mundial, encara que el nombre d’individus que la pateixen augmenta ràpidament, especialment en els països desenvolupats. Es l’alteració metabòlica més freqüent entre els humans i comporta també l’aparició de complicacions secundàries la retinopatia, neuropatia a més d’alteracions cardiovasculars. En un estudi poblacional realitzat pel Consell Assessor sobre la Diabetis de Catalunya (1997) s´observa que la prevalència de la DM coneguda a Catalunya és del 6,7 % i que augmenta en relació amb l´edat , de manera que la prevalència de la DM-2 global ( coneguda i no coneguada) és del 10,3% en la població de 30 a 89 anys (19).
En l’actualitat, existeixen teràpies pels dos tipus de diabetis que resulten insatisfactòries perquè no ofereixen una cura de la malaltia i en la majoria dels casos, no poden evitar l’aparició de les complicacions secundàries associades a la mateixa. El transplantament d’illots pancreàtics ha estat, sens dubte, una encoratjadora estratègia per a restaurar la massa de cèl.lula ß funcional en els pacients diabètics i poder així aconseguir la normoglicèmia; no obstant presenta limitacions importants: rebuig de empel i el gran nombre de pàncreas necessaris per a l’obtenció d’un nombre òptim d’illots (almenys dos donants/pacients) (20). Aixó comporta la necessitat d’ identificar noves teràpies gèniques i/o cel.lulars com per exemple l’obtenció de cèl.lules productores d’insulina a partir de cèl.lules pluripotents. Per fer-ho, és necessari aprofondir en els mecanismes moleculares de la propia cèl.lula beta que s’intenta reestablir. La viabilitat d’aquesta nova estratègia cèl.lular depèn principalment de tres importants prerequisits:
• Identificació d’unes cèl.lules pluripotencials o unes cèl.lules progenitores pancreàtiques que tinguin la capacitat d’autoreplicar-se i generar cèl.lules diferenciades.
• Identificació dels senyals proliferatius que permetin expandir d’una manera específica aquests progenitors pancreàtics.
• Identificació de senyals instructius que indueixin la diferenciació d’aquestes cèl.lules pluripotencials o progenitores en cèl.lules ß funcionals que secreten l´insulina correctament processada, d’una manera pulsàtil, en resposta a concentracions fisiològiques de glucosa.
A diferència de cèl-lules que provenen del mesoderm (cardiomiòcits) o les que provenen de l´ectoderm (cèl.lules neuronals), que poden ser diferencides espontàniament in vitro, les cèl.lules derivades de l´endoderm entre les que s´inclouen les cèl.lules Beta pancreàtiques, requereixen la presència de factors generalment desconeguts que són els que dirigieixen la diferenciació cel.lular.
Existeix bàsicament, dues estrategies per obtenir cèl.lules Beta a partir de cèl.lules pluripotencials embrióniques:
1.- Estratègia de Trapping, utilitzada per l’ equip del Dr.B. Soria. En aquests cas, cèl.lules mare són transfectades amb una construcció que porta un gen de resistencia a antibiòtic ( neomicina ) que està sota el control del gen de la insulina. D’aquesta manera, es seleccionen les cèl.lules que al diferenciar-se activaran únicament aquest promotor. Posteriorment s´afagí a la construcció un marcador no tòxic, però fàcilment perceptible ( la proteina fluorecent verde o GFP) , de forma que les cèl.lules co-expressen insulina i proteina verda , permeten una selecció més acurada de les maiteixes. D´aquesta manera s´establí l’èquipt del Dr. Soria, establí la linea cèl.lular IB/3x-99 derivada directament d´un conreu de cèl-lules ES de ratolí. Aquestes cèl-lules tenen la capacitat de formar agregats cèl.lular secretors d’insulina de manera glucosa depenent (21).
Per provar la capacita funcional d´aquestes cèl-lules “in vivo”, els agregats cèl-lulars foren implantats en la melsa de ratolins. La majoria dels animals van controlar la glucèmia, encara que el 40% de ratolins trasplantats desenvoluparen una hiperglucèmia a les 12 setmanes (22). Així doncs, l´estudi estableix encara clares dificultats.
Varies raons poden explicar la pèrdua de les cèl.lules trasplantades:
1.- pèrdua de les característiques genotípiques en absència de la pressió selectiva.
2.- Mort induïda de la cèl-lula , potser deguda al nou nínxol cèl-lular.
3.- Rebuig per part de l´hoste a l´empelt cèl-lular.
4.- Producció de tumors.
2.- Marcadors selectius de cèl.lules mare pancreàtiques
La Nestina és una proteina del filament , que s´ha identificat per ser un marcador de cèl.lules Stem del Sistema Nerviós Central (SNC).Donat que existiesen moltes similituds entre el SNC i el pàncrees,Lumelsky et al , proposaren la nestina com un marcador específic de la cèl-lula stewm endocrina (23). El protocol emprat per aquests investigadors, va demostrar que després la deprivació en el medi de conreu de LIF (Leukimia factor inhibitor) o suplementant aquests amb bFGF, permetia la formació d´agregats cèl-lulars que expresan, no només insulina sinó tambè glucagó, somatostatina i polpèptid pancreàtic, hormones que es troben en els illots de Langerhans. No s´identificà cap marcador exocrí com es l’amilasa o la carboxypeptidase A. Anàlisi histològics demostraren que els “clusters” cel.lulars tenien característiques comunes amb l´estructura normal de l´illot, però únicament un 50 % de les cèl-lules expresavan insulina.
Estudis funcionals de secreció, van donar mostres de resposta de manera dosi-depenent.
L’estratègia de seleccionar cèl.lules que expressen la proteïna Nestina ha estat criticada, encara que existiesen evidencies clares de que pot ser un bon marcador.
Existeix una superposició en els factors de transcripció de la neurogènesi i l´endocrinogènesi, així com factors claus neuronals del bHLH, com neurogenin-1, 2, NeuroD i NeuroD2 poden ajudar a activar el gen de l´insulina en una cèl.lula neuronal, i són factors idèntics (NeuroD), o molt similar (Ngn1 o 2) a aquests components del bHLH necessaris per la funció i desenvolupament de la cèl.lula endocrina (NeuroD y Ngn3) (24,25).
No obstant , el marcador específic endodermal HNF3b, s´expressa en cèl.lules derivades de les embriòniques , a més d´altres marcadors han de ser explorat per poder compendre el procés íntim de com una cèl-lula indiferenciada es trasforma en una cèl-lula amb fenotipus beta.
En el cas del pàncrees adult no hi ha consens sobre quin tipus cel.lular podria ser la verdadera Stem Cell pancreàtica (26). S’ha descrit que en el pàncreas existeix un procés de neogènesis; és a dir, de formació ex novo d’illots pancreàtics, que augmenta en diferents situacions tant en humans (27) com en models animals (28-30). Alguns autors postulen que després d’una pancreatectomia parcial, algunes cèl.lules ductals madures podrien començar a proliferar i desdiferenciar-se fins a donar una cèl.lula amb capacitat multipotencial, per poder diferenciar-se en cèl.lules de l’illot, cèl.lula acinar o cèl.lula ductal madura; la direcció de diferenciació d’aquelles cèl.lules dependria de senyals externes o morfògenes (31). En aquest cas parlaríem de Cèl.lules mare funcionals de manera semblant al que s’ha descrit en el fetge i en altres teixits (32-34). Altres autors han descrit l’existència de possibles Cèl.lules mare en els mateixos illots pancreàtics (35-36) i han suggerit que aquestes són una font de noves cèl.lules de l’illot. Per tant, encara que els experiments han demostrat que en el pàncreas existeix un procés regeneratiu tant intra-illot com extra-illot, la detecció i caracterització de les Cèl.lules mare pancreàtiques està resultant un feina difícil. La falta d’uns marcadors clars (37-39) així com d’assatjos fiables, ha obstaculitzat aquesta dificultosa tasca. No obstant, sabem que noves cèl.lules Beta es generen durant la vida adulta i que, en part, provenen de cèl.lules que es troben en el duct pancreàtic. Una rigorosa caracterització del procés del desenvolupament embrionari del pàncrees, dels mecanismes moleculars de proliferació i apoptosi dels illots pancreàtics, així com dels possibles candidats a Cèl.lules mare descrits ens hauria de permetre aillar les Cèl.lules mare adultes pancreàtiques. La possibilitat d’ expansió i diferenciació d’aquestes cèl.lules ens permet obrir l’esperança d’obtenir suficients cèl.lules Beta per la teràpia cel.lular. Encara més apassionant és la possibilitat de que, a partir d’una petita mostra de teixit pancreàtic del pacient, es poguesin obtenir in vitro nous illots que es transplantarien posteriorment al paciente. D’aquesta manera els problemes de rebuig ja no existirien.
Com a conclusió podriam dir que ens trobem en l’era d’obrir nou camp en l’ingeneria cel.lular, que segons definició feta pel Dr. Bernat Soria es:“un nou camp interdisciplinar que aplica els principis de l’ ingenieria i de les ciències de la vida a l´obtenció de substituts biològics per restaurar, mantenir o millorar la funció tisular”.
1. Peter J Donovan and John Gearhart (2001). The end of the beginning for pluripotent Stem Cells. Nature 414: 92-97.
2. Martin G (1981). Isolation of a Pluripotent Cell Line from Early Mouse Embrios Cultured in Medium Conditioned by Teratocarcinoma Cells. Proc. Natl. Acad. Sci. USA 78: 7634-7638.
3. Evans MJ, Kaufman MH (1981). Establishment in Culture of Pluripotential Stem Cells from Mouse Embrios. Nature 291: 154-156.
4. Thomson JA, Itskovitz-Eldor J, Shapiro SS, I colbs. (1998). Embrionic Stem Cell Lines Derived from Human Blastocits. Science 282: 1145-1147.
5. Reubinoff BE, Pera MF, Fong CI, Trouson A, Bongso A (2000). Embrionic Stem Cells Lines from Human Blastocists: Somatic Differentation in vitro. Nature Biotechol. 18: 399-404.
6. Bishop AE, Butteri LDK, Polak JM (2002) Embrionic Stem Cells. J. Pathol. 197: 424-429.
7. Mezey E, Chandross KJ (2000). Bone marrow: a possible alternative source of cells in the adult nervous system. Eur J Pharmacol 405(1-3):297-302.
8. Poulsom R, Alison MR, Forbes SJ, Wright NA (2002). Adult Stem Cell plasticity. J Pathol 197:441-456.
9. Björklund A and Svendsen CN (2001). Chimeric Stem Cells. Trends in Molecular Medicine 7: 144-146.
10. Blackburn EH, Szostak JW (1984). Molecular structure of centromeres and telomereses. Ann Rev Biochem 53:163-94.
11. Petersen BE i colbs. (1999). Bone marrow as a potential source of hepatic oval cells. Science 284:1168-70.
12. Alison MR i colbs. (2000). Hepatocyte from non-hepatic adult Stem Cells. Nature 406, 257-260.
13. Ferrari G i colbs. (1998). Muscle regeneration by bone marrow-derived myogenic progenitors. Science 279:1528-1530.
14. Ying QL, Nichols J, Evans EP, Smith AG. (2002). Changing Potency by spontaneous fusion. Nature 4;416(6880).
15. Vescovi AL, Rietze R, Magli MC, Bjornson C (2002). Hematopoietic potential of neural Stem Cells. Nat Med. 2002 Jun;8(6):535; author reply 536-7.
16. Wagers AJ, Sherwood RI, Christensen JL, Weissman IL (2002). Little evidence for developmental plasticity of adult hematopoietic Stem Cells. Science. 27; 297(5590):2256-9.
17. Hori J, Tat Fong Ng, Shatos M, Klassen H, Streilein JW, Young MJ (2003). Neural Progenitor Cells Lack Immunogenicity and Resist Destruction as Allografts. Stem Cells 21:405-416.
18. Liechty KW, MacKenzie TC, Shaaban AF, Radu A, Moseley AM, Deans R, Marshak DR, Flake AW (2000). Human mesenchymal Stem Cells engraft and demonstrate site-specific differentiation after in utero transplantation in sheep. Nat Med.6(11):1282-6.
19. Generalitat de Catalunya, departament de Sanitat i Seguretat Social.(1997) Prevalença de diabetis Mellitus no insulinodependent a catalunya. Bulletí Epidemiològic de catalunya 3.41-43.
20. Shapiro AM, Lakei JR, Rian EA, i colbs. (2000). Islet Transplantarion in Seven Patients with Type I Diabetes Mellitus Using a Glucocorticoid-Free Immunosuppressive Regimen. N. Engl. J. Med. 27: 230-238.
21. Soria B, Skoudy A, Martin F (2001). From Stem Cells to beta cells: new strategies in cell therapy of diabetes Mellitus. Diabetologia 44(4): 407-415.
22. Soria B, Roche E, Berna G, Leon-Quinto T, Reig JA, Martin F.(2000) Insulin-secreting cells derived from embryonic Stem Cells normalize glycemia in streptozotocin-induced diabetic mice. Diabetes 49(2):157-162.
23. Lumelsky N, Blondel O, Laeng P, Velasco I, Ravin R, McKay R.(2001) Differentiation of embryonic Stem Cells to insulin-secreting structures similar to pancreatic islets. Science 92 (5520):1389-1394.
24. Jenny M, Uhl C, Roche C, Duluc I, Guillermin V, Guillemot F, Jensen J, Kedinger M, Gradwohl (2002). Neurogenin3 is differentially required for endocrine cell fate specification in the intestinal and gastric epithelium. G.EMBO J. 2;21(23):6338-47.
25. Gallo R, Zazzeroni F, Alesse E, i colbs. (2002). REN: a novel, developmentally regulated gene that promotes neural cell differentiation. J Cell Biol 19;158(4):731-40.
26. Bonner-Weir S, Sharma A (2002). Pancreatic Stem Cells. J. Pathol 197: 519-526.
27. Gepts W (1965). Pathological Anatomy of the Pancreas in Juvenile Diabetes. Diabetes 14: 619-633.
28. Weaver CV, Sorenson RL, Kaung HC (1985). Immunocitochemical Localization of Insulin-Immunoreactive Cells in the Ducts of Rats Treated with Tripsin Inhibitor. Diabetologia 28:0781-785.
29. Bonner-Weir S, Baxter LA, Schuppin GT, Smith FE (1993). A Second Pathwai for Regeneration of the Adult Exocrine and Endocrine Pancreas: a Possible Recapitulation of Embrionic Development. Diabetes 42: 1715-1720.
30. Xu G, Stoffers DA, Habener JF, Bonner-Weir S (1999). Exendin-4 Stimulates both ß-Cell Replication and Neogenesis, Resulting in Increased ß-Cell Mass and Improved Glucose Tolerance in Diabetic Rats. Diabetes 48: 4909-4918.
31. Sharma A, Zangen DH, Reitz P, I colbs. (1999). The Homeodomain Protein IDX-1 Increases after a Earli Burst of Proliferation During Pancreas Regeneration. Diabetes 48: 507-513.
32. Potten C (1998). Stem Cells in Gastrointestinal Epithelium: Numbers, Characteristics and Death. Phil. Trans. R. Soc. Lond. B. 353: 821-830.
33. Alison M, Sarraf C (1998). Hepatic Stem Cells. J. Hepatol. 29: 678-683.
34. Block GD, Locker J, Bowen WC, I colbs. (1996). Population Expansion, Clonal Growth, and Specific Differentiation Patterns in Primary Culture of Hepatocites Induced bi HGF/SF, EGF and TGFß in a Chemicalli Defined (HGM) medium. J. Cell. Biol. 132: 1133-1149.
35. Guz I, Nasir I, Teitelman G (2001). Regeneration of Pancreatic ß Cells from Intra-Islet Precursor Cells in an Experimental Model of Diabetes. Endocrionologi. 142: 4956-4968.
36. Zulewski H, Abraham EJ, Gerlach MJU, I colbs. (2001). Multipustential Nestin-Positive Stem Cells Isolated from Adult Pancreatic Islets Differentiate Ex Vivo into Pancreatic Endocrine, Exocrine and Hepatic Phenotipes. Diabetes 50: 521-533.
37. Bouwens L, Wang RN, De Bali E, Pipeleers DG, Kloppel G (1994). Citokeratins as Markers of Ductal Cell Differentiation and Isle Neogenesis in the Neonatal Rat Pancreas. Diabetes 43: 1279-1283.
38. Offield MF, Jetton TL, Laboski P, i colbs. (1996). PDX-1 Is Required for Pancreatic Outgrowth and Differentiation of the Rostral Duodenum. Development 122: 983-985.
39. Teitelman G, Alpert S, Polak JM, Martinez A, Hanahan D (1993). Precursor Cells of Mouse Endocrine Pancreas Copexpress Insulin, Glucagon and the Neuronal Proteins Tirosine Hidroxilese and Neuropeptide I but not Pancreatic Polipeptide. Development 118:1031-1039.
Fig.1 Origen de les cèl.lules mare d’origen embriònic. Esquema reproduit de la publicació. The end of the beginning for pluripotent Stem Cells. Nature 414: 92-97.
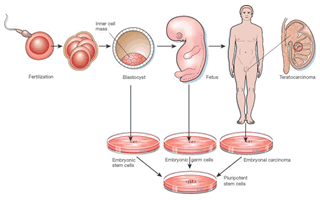
Fig.2 Esquema on has representa els posibles origens de les cèl.lules mare adultes. Reproducció extreta de la publicació. Trends Mol Med (2001)7(4). 144-6